{kind=link}
{kind=link}
{kind=link}
Ett sömnproblem kan uppstå i flera regleringsnivåer. Sömnen kan vara för kort, ligga fel i dygnsrytmen, vara fragmenterad, sakna återhämtande kvalitet eller störas av överaktiva vakenhetssystem. Samma person kan ha flera mekanismer samtidigt.
En användbar grundmodell är att skilja mellan sömnstadier, sömntryck, dygnsrytm och vakenhetssystem. Den modellen gör det lättare att förstå varför kaffe, skärmljus, dagsömn, alkohol, centralstimulantia, ångest, sömnapné och sömnläkemedel påverkar sömnen på olika sätt.
Sömnstadier och sömnarkitektur
Indelningen i sömnstadier bygger på polysomnografi. Då registreras bland annat hjärnans elektriska aktivitet med EEG, ögonrörelser med EOG, muskeltonus med EMG, andning, syremättnad, puls och kroppsrörelser. I klinisk vardag används inte polysomnografi för alla sömnproblem, men sömnstadierna ger ett språk för skillnaden mellan lätt sömn, djupsömn och REM-sömn.
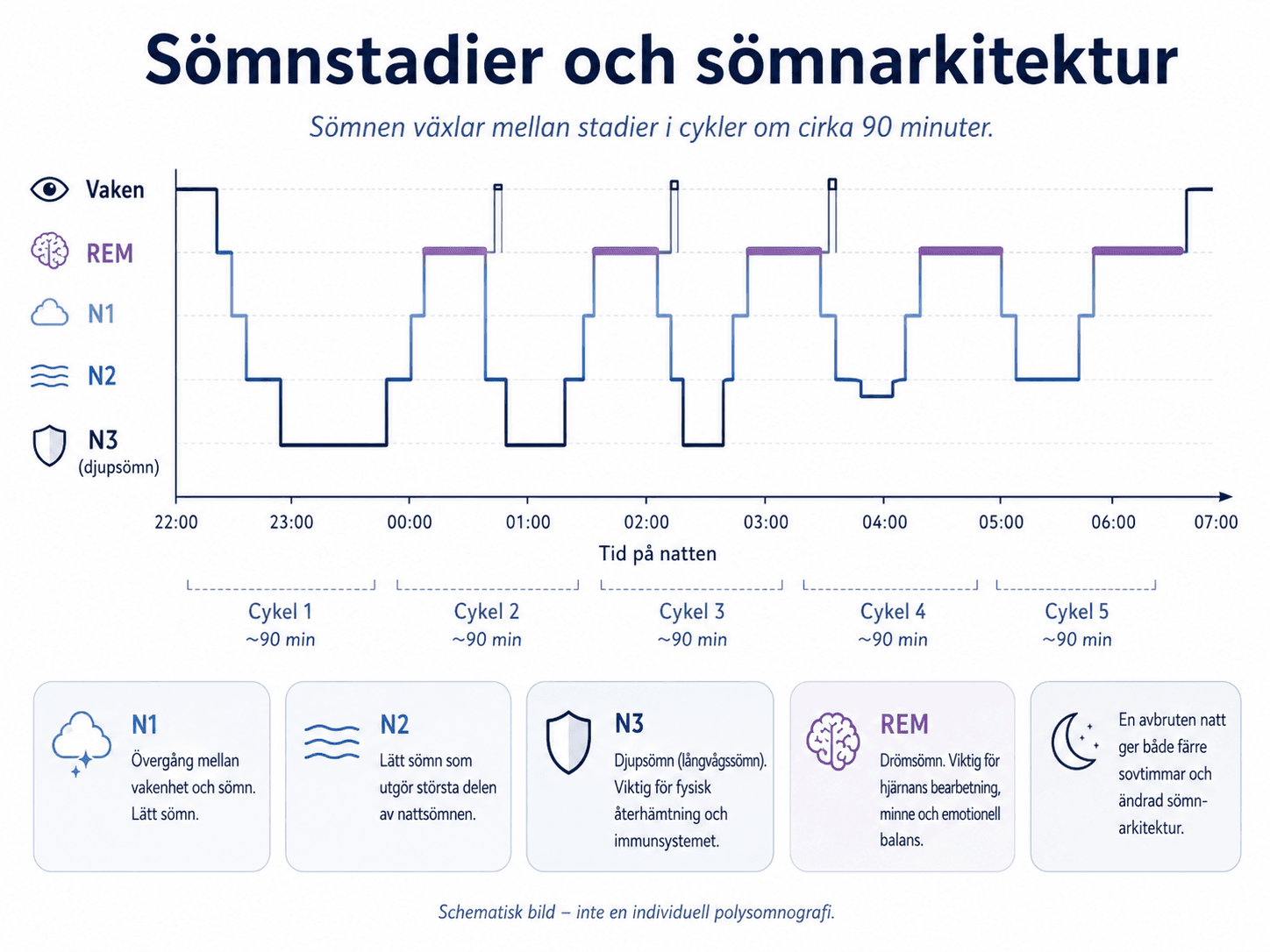
Människans sömn delas in i NREM-sömn och REM-sömn. NREM-sömnen delas i N1, N2 och N3.
N1 är övergången mellan vakenhet och sömn. Den är lättstörd. Personen kan uppleva att hen inte har sovit trots att sömn registreras.
N2 utgör ofta den största delen av nattsömnen hos vuxna. EEG visar bland annat sömnspolar och K-komplex. N2 är stabilare än N1 men räknas inte som djupsömn.
N3 motsvarar djupsömn eller slow-wave sleep. Den kännetecknas av långsamma EEG-vågor och högre väckningströskel. Djupsömn förekommer vanligen mest under första delen av natten.
REM-sömn kännetecknas av snabba ögonrörelser, låg muskeltonus och hög hjärnaktivitet. Livliga drömmar är vanligare under REM-sömn, även om drömupplevelser kan förekomma i andra sömnstadier.
En normal natt består av upprepade cykler mellan NREM och REM. En cykel varar ofta omkring 90 minuter, med individuell variation. Första delen av natten innehåller vanligen mer N3. Sista delen innehåller vanligen mer REM. En natt som avbryts tidigt innebär därför både färre sovtimmar och förändrad sömnstadiesammansättning.
Sömnarkitekturen påverkas av ålder, alkohol, läkemedel, depression, smärta, sömnapné, neurologiska tillstånd, abstinens och dygnsrytm. Alkohol kan förkorta insomningstiden men fragmenterar ofta sömnen senare under natten. Sederande läkemedel kan göra personen trött utan att sömnen får normal struktur.
Homeostatisk sömnreglering: process S
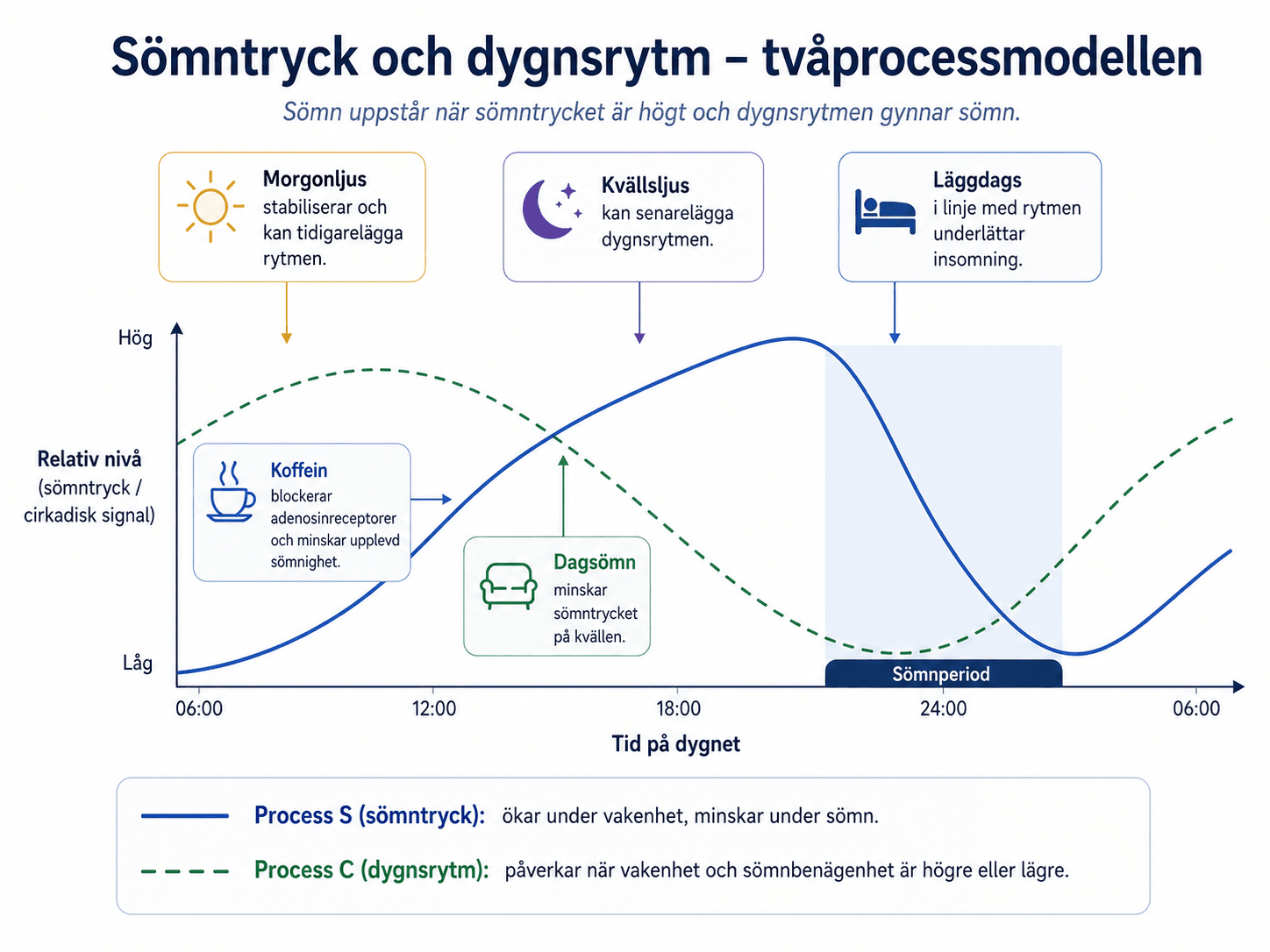
Den homeostatiska sömnregleringen beskriver hur sömnbenägenheten ökar med tiden vaken. I tvåprocessmodellen kallas detta process S. Under sömn minskar sömntrycket igen.
Djupsömn och långsamvågsaktivitet i EEG används ofta som markörer för homeostatisk sömnreglering. Efter sömnbrist ses vanligen ökad långsamvågsaktivitet när personen får sova.
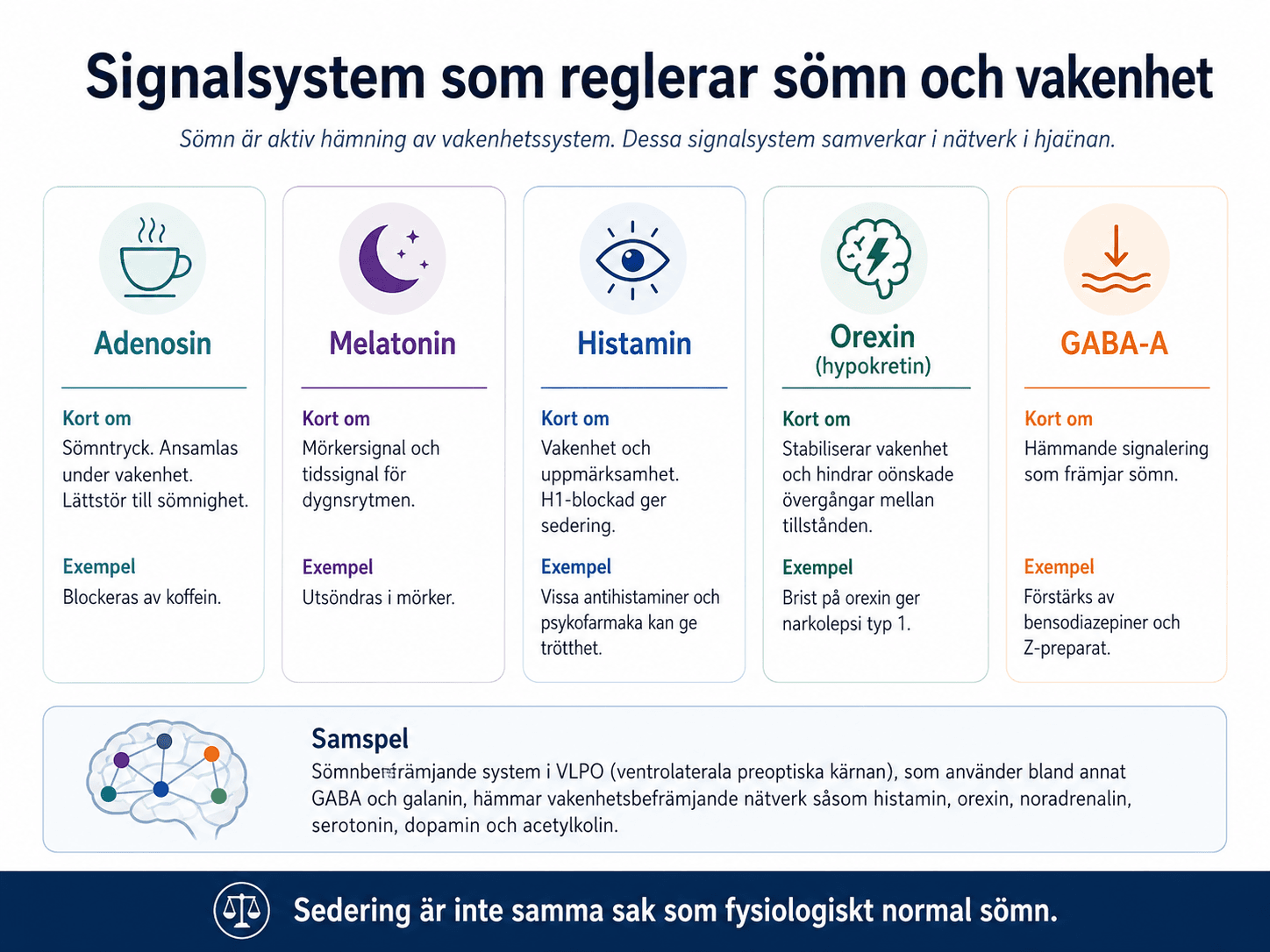
Adenosin är en av de signalmolekyler som kopplas till sömntryck. Adenosin ansamlas under vakenhet och bidrar till sömnighet. Koffein blockerar adenosinreceptorer och minskar den upplevda sömnigheten. Koffein minskar inte sömnbehovet.
Mycket tid i sängen dagtid, lång dagsömn och låg aktivitetsnivå kan minska sömntrycket på kvällen. Koffein sent kan göra att personen inte känner sömntrycket trots att sömnbehovet finns kvar.
Cirkadisk reglering: process C
Dygnsrytmen beskriver kroppens ungefärliga 24-timmarsvariation i sömnbenägenhet, vakenhet, kroppstemperatur, hormoner, ämnesomsättning och prestationsförmåga. I tvåprocessmodellen kallas den cirkadiska komponenten process C.
Den centrala biologiska klockan sitter i suprachiasmatiska kärnan i hypotalamus. Den får ljusinformation från näthinnan och synkroniserar kroppens rytmer med omgivningens ljus–mörkercykel. Ljus på morgonen tenderar att tidigarelägga rytmen. Ljus sent på kvällen eller natten tenderar att senarelägga rytmen.
Melatonin utsöndras under mörker och används som markör för cirkadisk fas. Dim light melatonin onset, DLMO, används i forskning och specialiserad klinik för att bedöma när kroppens biologiska kväll börjar. Melatonin är främst en tidssignal, inte ett allmänt sövande medel i samma mening som GABA-verkande hypnotika.
Process S och process C kan dra åt olika håll. En person kan ha högt sömntryck men ändå svårt att somna om den biologiska rytmen ligger sent. En annan kan ha låg sömnbenägenhet på kvällen efter lång dagsömn trots att klockan visar läggdags.
Vakenhetssystem
Vakenhet är ett aktivt neurobiologiskt tillstånd. Den hålls uppe av nätverk i hjärnstam, hypotalamus och basala framhjärnan. Viktiga signalsubstanser är histamin, noradrenalin, serotonin, dopamin, acetylkolin och orexin/hypokretin.
Histaminerga neuron i tuberomamillära kärnan i hypotalamus bidrar till vakenhet. Blockad av centrala H1-receptorer ger sedering. Det förklarar varför vissa antihistaminer, och även läkemedel med stark H1-blockad som vissa antidepressiva och antipsykotika, kan ge trötthet.
Orexin, även kallat hypokretin, produceras i laterala hypotalamus. Systemet stabiliserar vakenhet och gränserna mellan vakenhet, NREM-sömn och REM-sömn. Brist på orexinsignalering är central vid narkolepsi typ 1.
Noradrenalin från locus coeruleus, serotonin från raphekärnor, dopamin från mesencefala system och acetylkolin från basala framhjärnan och hjärnstam bidrar till uppmärksamhet, aktivering, motivation, REM-reglering och kortikal aktivitet. Psykofarmaka, centralstimulantia, nikotin och droger kan påverka dessa system.
När vakenhetssystemen dominerar kan hög trötthet samexistera med oförmåga att somna. Det ses till exempel vid ångest, abstinens, centralstimulantiapåverkan, smärta och uppvarvning.
Sömnfrämjande system
Sömn kräver aktiv hämning av vakenhetssystem. Neuron i ventrolaterala preoptiska området i hypotalamus använder bland annat GABA och galanin för att hämma vakenhetsbefrämjande nätverk.
Samspelet mellan sömnfrämjande och vakenhetsfrämjande nätverk beskrivs ofta som en flip-flop-modell. När vakenhetssystemen dominerar hålls hjärnan vaken. När sömnfrämjande system dominerar stabiliseras sömn. Orexin bidrar till att stabilisera läget och minska oönskade övergångar mellan tillstånden.
Detta förklarar varför sömn inte alltid kommer gradvis. En person kan ligga vaken länge och sedan somna snabbt när aktiveringsnivån sjunker. Om vakenhetssystemen fortsätter dominera kan personen vara utmattad men ändå inte somna.
REM-reglering
REM-sömn har särskild reglering. Hjärnan är aktiv, ögonrörelserna snabba och skelettmuskulaturen hämmas normalt. Denna REM-atoni hindrar att dröminnehåll omsätts i större rörelser.
Instabil REM-reglering kan ge REM-fenomen i övergången mellan sömn och vakenhet. Sömnparalys innebär uppvaknande med kvarstående muskelatoni. Hypnagoga och hypnopompa hallucinationer kan förekomma vid insomning eller uppvaknande. Kataplexi är plötslig muskelsvaghet utlöst av känslor och är starkt kopplad till narkolepsi typ 1.
Vid REM sleep behavior disorder saknas normal REM-atoni, vilket kan leda till att personen agerar ut drömmar. Tillståndet är särskilt viktigt hos äldre eftersom det kan vara kopplat till neurodegenerativ sjukdom.
Substanser och läkemedel som visar systemen
Koffein visar adenosinsystemet. Det blockerar adenosinreceptorer och minskar sömnighet utan att minska sömnbehovet.
Nikotin visar kolinerg aktivering. Det kan bidra till insomningssvårigheter, fragmenterad sömn och tidigt uppvaknande. Nikotinabstinens kan också störa sömnen.
Alkohol visar skillnaden mellan sedering och sömnkvalitet. Alkohol kan göra insomningen lättare men ger ofta mer fragmenterad sömn senare under natten och kan förvärra snarkning och obstruktiv sömnapné.
Antihistaminer visar histaminsystemet. H1-blockad ger sedering, men sedering är inte samma sak som fysiologiskt normal sömn.
Bensodiazepiner och Z-preparat visar GABA-systemet. De förstärker GABA-A-signalering och kan minska insomningstid, men påverkar även minne, reaktionsförmåga, sömnarkitektur, tolerans och beroenderisk.
Melatonin visar dygnsrytmens timing. Effekten beror i hög grad på när preparatet tas i förhållande till personens biologiska rytm.
Läkemedelsval, dosering och utsättning kräver medicinsk bedömning.
Fyra signalsystem som ofta syns i vardagen
Adenosin: sömntryck. Blockeras av koffein.
Melatonin: mörkersignal och markör för dygnsrytmens timing.
Histamin: vakenhet. H1-blockad ger sedering.
Orexin: stabiliserar vakenhet. Brist ger narkolepsi typ 1.
GABA: hämmande signalering. Förstärks av bensodiazepiner och Z-preparat.
Fall
Rami är 39 år och har tidigare centralstimulantiaberoende, ångest och perioder med depression. Han säger att han vill sova tidigare men somnar sällan före 03. Han dricker energidryck på kvällen eftersom han annars “kraschar” och blir nedstämd. Han ligger ofta i sängen från 22 men använder telefonen i flera timmar. Dagtid sover han ibland två timmar efter lunch.
Mönstret kan beskrivas med fysiologiska mekanismer. Dagsömnen minskar sömntrycket på kvällen. Energidryck blockerar adenosinsignalering. Ljus och skärm sent kan senarelägga dygnsrytmen. Ångest och tidigare centralstimulantiabruk kan hålla vakenhetssystemen aktiverade. Tiden i sängen överskattar den faktiska sömnen.
Ramis önskemål om “något starkt att sova på” fångar inte mekanismerna. Sömntryck, dygnsrytm, adenosin/koffein, aktiveringsnivå, substanshistorik och faktisk sömntid behöver hållas isär. Om läkemedel övervägs behöver beroenderisk, aktuell substansstatus, psykisk instabilitet och möjlig sömnstörning bedömas medicinskt.
Referenser
- Troester MM, Quan SF, Berry RB; American Academy of Sleep Medicine. The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications. Version 3. Darien, IL: American Academy of Sleep Medicine; 2023.
- Borbély AA. A two process model of sleep regulation. Human Neurobiology. 1982;1(3):195–204.
- Dijk DJ, Czeisler CA. Contribution of the circadian pacemaker and the sleep homeostat to sleep propensity, sleep structure, electroencephalographic slow waves, and sleep spindle activity in humans. Journal of Neuroscience. 1995;15(5):3526–3538.
- Scammell TE, Arrigoni E, Lipton JO. Neural circuitry of wakefulness and sleep. Neuron. 2017;93(4):747–765.
- Saper CB, Fuller PM, Pedersen NP, Lu J, Scammell TE. Sleep state switching. Neuron. 2010;68(6):1023–1042.
- Brown RE, Basheer R, McKenna JT, Strecker RE, McCarley RW. Control of sleep and wakefulness. Physiological Reviews. 2012;92(3):1087–1187.
- Reichert CF, Deboer T, Landolt HP. Adenosine, caffeine, and sleep–wake regulation: state of the science and perspectives. Journal of Sleep Research. 2022;31(4):e13597.
- Haas HL, Sergeeva OA, Selbach O. Histamine in the nervous system. Physiological Reviews. 2008;88(3):1183–1241.
- Sakurai T. The role of orexin in motivated behaviours. Nature Reviews Neuroscience. 2014;15(11):719–731.
- Arendt J, Skene DJ. Melatonin as a chronobiotic. Sleep Medicine Reviews. 2005;9(1):25–39.